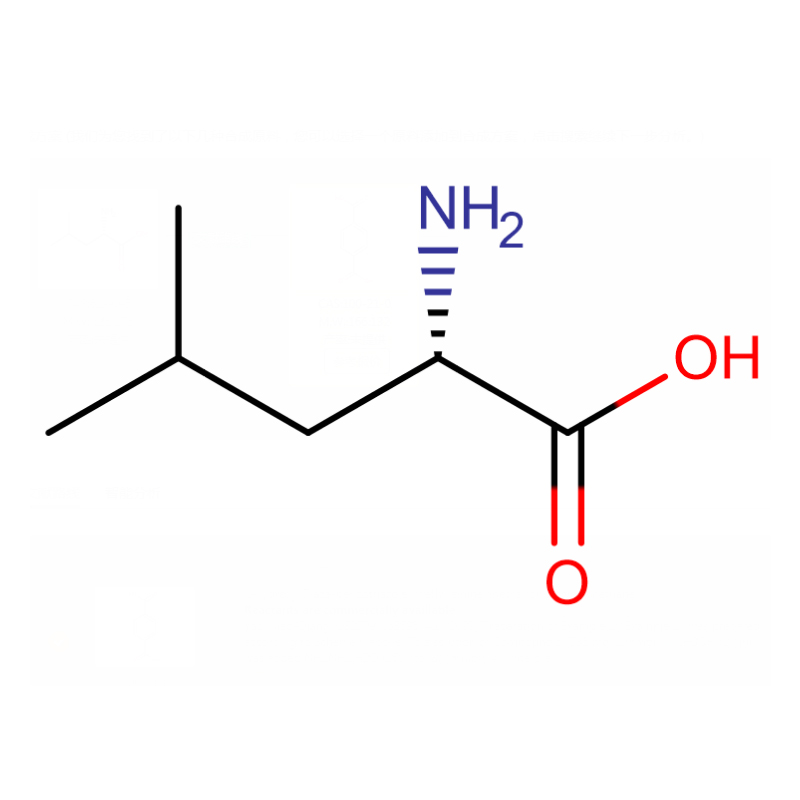
L-leutsiin Cas: 61-90-5 99% valge pulber
| Kataloogi number | XD90304 |
| tootenimi | L-leutsiin |
| CAS | 61-90-5 |
| Molekulaarvalem | C6H13NO2 |
| Molekulmass | 131.17292 |
| Salvestusruumi üksikasjad | Ambient |
| Harmoneeritud tariifikoodeks | 29224985 |
Toote spetsifikatsioon
Sarnaselt teistele positiivse ahelaga RNA viirustele replitseeritakse tombusviiruseid membraaniga seotud viiruse replikaasi kompleks (VRC).VRC koosneb p92 viirusest kodeeritud RNA-sõltuvast RNA polümeraasist (RdRp), viiruse p33 RNA chaperoonist ja mitmest koopteeritud peremeesvalgust.Selleks, et saada pärast translatsiooni funktsionaalne RdRp, tuleks p92 replikatsioonivalk lisada VRC-sse, millele järgneb selle aktiveerimine.Oleme varem näidanud rakuvaba pärmiekstraktil põhinevas testis, et Tomato bushy stunt virus (TBSV) RdRp aktiveerimine nõuab lahustuvat peremeesfaktorit.Selles artiklis tuvastame rakulise kuumašoki valgu 70 (Hsp70) kui koopteeritud peremeesfaktorit, mis on vajalik N-terminaalselt kärbitud rekombinantse TBSV RdRp aktiveerimiseks.Lisaks pärsib Hsp70 funktsiooni väikesemolekuliline blokeerimine RNA sünteesi tombusviiruse RdRp poolt in vitro.Lisaks näitame, et neutraalsed fosfolipiidid, nimelt fosfatidüületanoolamiin (PE) ja fosfatidüülkoliin (PC ), suurendavad RdRp aktivatsiooni in vitro .Seevastu fosfatidüülglütserool (PG) avaldab tugevat ja domineerivat inhibeerivat toimet in vitro RdRp aktivatsioonile.Samuti demonstreerime, et PE ja PC stimuleerivad RdRp-viiruse plussahela RNA [(+)RNA] interaktsiooni, samas kui PG inhibeerib viiruse RNA seondumist RdRp-ga.Tuginedes erinevate fosfolipiidide stimuleerivatele ja inhibeerivatele rollidele tombusviiruse RdRp aktiveerimisel, teeme ettepaneku, et tombusviirused võivad kasutada sihtmärgiks olevate rakuväliste membraanide lipiidide koostist, et reguleerida uut VRC koostu infektsiooni käigus. Viiruse poolt kodeeritud RNA-st sõltuv RNA polümeraas (RdRp), mis vastutab viiruse RNA järglaste sünteesi eest mitme positiivse ahelaga RNA viiruse nakatunud rakkudes, on esialgu inaktiivne.See strateegia väldib tõenäoliselt viiruse RNA sünteesi tsütosoolis, mis viiks kiiresti RNA-ga vallandatud raku viirusevastaste vastuste esilekutsumiseni.Membraaniga seotud replikaasikompleksi kokkupanemise ajal aktiveerub viiruse RdRp mittetäielikult mõistetava protsessi kaudu, mis muudab RdRp võimeliseks RNA sünteesiks.Kasutades TBSV RdRp-d, näitame, et koopteeritud rakuline Hsp70 chaperone ja neutraalsed fosfolipiidid hõlbustavad RdRp aktiveerimist in vitro.Seevastu fosfatidüülglütseroolil (PG) on domineeriv inhibeeriv toime in vitro RdRp aktivatsioonile ja RdRp-viiruse RNA interaktsioonile, mis viitab sellele, et RdRp-d ümbritsev membraanne mikrodomeen mõjutab suuresti selle võimet RNA sünteesiks.Seega sõltub viiruse RdRp aktiveerimine tõenäoliselt mitmest peremeeskomponendist nakatunud rakkudes.
4-nitrofenüül-N-atsetüül-β-D-glükosaminiid on kasulik substraat N-atsetüül-b-glükosaminidaasi aktiivsuse kiireks kolorimeetriliseks testiks inimese uriinis.Kromogeenne β-glükoosaminidaasi substraat, mis annab lõhustamisel kollase lahuse, mida kasutatakse eriti pärm- ja hallitusseentes.